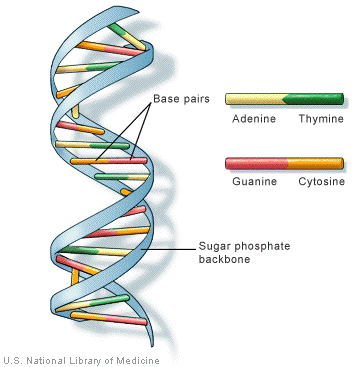
Mutation: Genetic recombination produces enormous number of variability for natural selection to choose from. But when one talks of millions of years of natural selection one should look for other mechanisms which would provide totally new factors and bring about fresh variability. Mutation provides totally new factors. In a broad sense a mutation is a hereditary change not due to the simple recombination of genes. Such genotypic changes include changes in chromosomal number, gross changes in the structure of chromosomes and changes in individual genes. Many mutations involve changes in single base pairs; this is called point mutation.

These mutations can be triggered by radiations and chemicals such as 5-Bromouracil, aminopurine, ethylmethane sulfonate etc. Mutations can take place in the reproductive or the non-reproductive cells of an organism. Those in the non-reproductive cells affect the host and are not passed on to its progenies and therefore are not important in evolution. Those mutations which occur in reproductive cells are passed on to progenies and are important to evolution.
Most mutations are lethal but some are beneficial. The earliest recorded mutation in domestic animals was that observed by Seth Wright in 1791 on his farm in Massachusetts. He noticed a peculiar male lamb with unusually short legs in his flock of sheep. It occurred to him that it would be advantageous to have a whole flock of these short-legged sheep which could not jump over the low stone fences. And so he used the new short-legged ram for breeding his fifteen ewes in the next season. Two of the fifteen lambs produced had short legs. Short-legged sheep were then bred together, and a line was developed in which the new trait was expressed in all individuals.

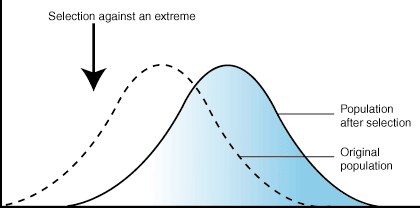
In this case the mutation was favourable to man and it is he who did the selection. Natural selection may or may not have favoured this mutation. Mutation alone cannot account for evolution; rather it furnishes the raw materials on which other forces act to bring about evolutionary change.